Convidado
GF Ouro
- Entrou
- Jun 2, 2010
- Mensagens
- 5,086
- Gostos Recebidos
- 0
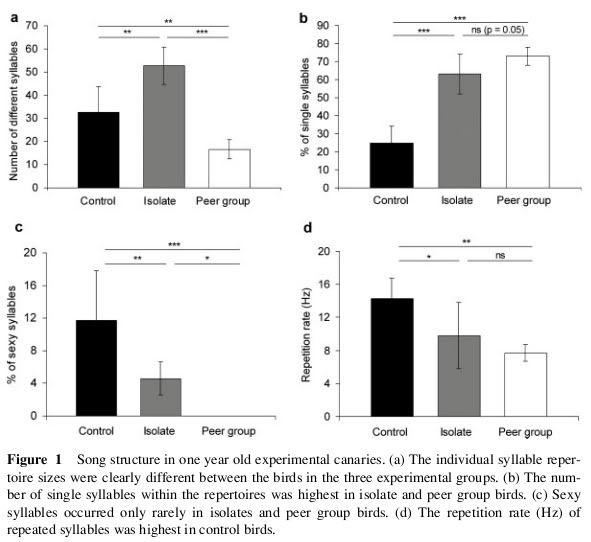
NEUROLÓGICAS CONTROLE DE APRENDIZAGEM
Embora o canto canário é executado em seu órgão melodiosa, a siringe, é o cérebro onde reside o conteúdo no "Stable Song", com sua composição silábica e padrões específicos musical e quando a sua execução é controlada pela a siringe, como discutido abaixo.
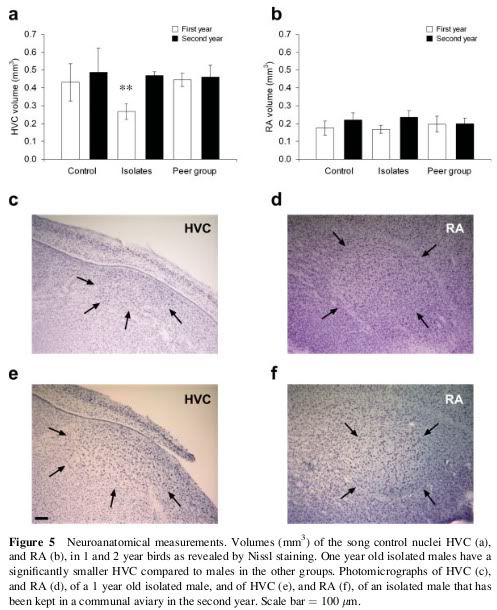
Em 1976, Fernando Nottebohm, um especialista em comportamento animal e neurogênese de The Rockefeller University, NY, E.U.A., identificados grupos distintos de células cerebrais no controle da cantando canários. Estas células são distribuídos em um grande núcleo, chamado de "maior centro vocal (CVS), que envia axonal outro núcleo denominado" centro robustus archistriatalis (RA), esta, por sua vez, envia outros para o núcleo de axonal " nervo craniano hipoglosso "que abastece os músculos da siringe.
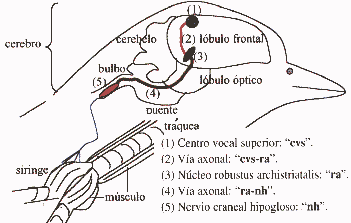
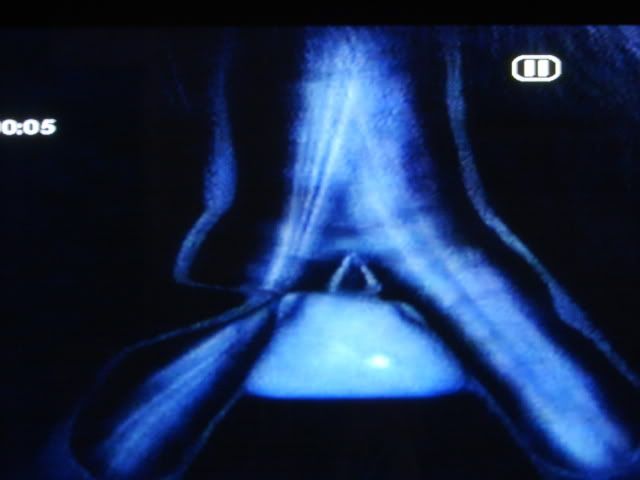
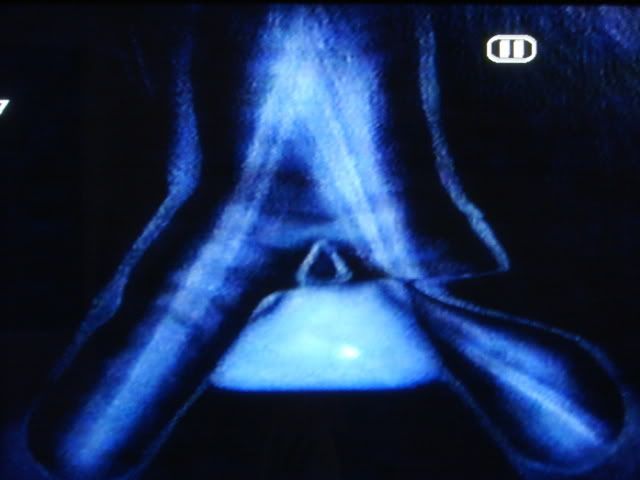
Cérebro Canárias. No desenho cima de um corte lateral do cérebro e na parte inferior do desenho de uma seção longitudinal da traquéia, brônquios e siringe do canário. O cérebro tem vários núcleos formados por neurônios especializados de controle de aprendizagem e performance da canção. Quanto maior o núcleo central superior vocal (CVS), que envia as extensões nervosas para o "núcleo Robustus archistriatalis" (M), que por sua vez, enviar outras extensões do nervo craniano ao núcleo do hipoglosso, que abastece os músculos da siringe: órgão onde o som é produzido. (Reproduzido de Nottebohm F. De canto dos pássaros a neurogênese. Sci Am 1989 Feb; 260 (2): 74-9).
Ambos os centros estão localizados nos lobos frontais das Canárias, que é onde você controla complexos comportamentos aprendidos. CVS parece ser o gerente da loja para identificar e produzir a música, na verdade, este kernel trabalha para reconhecer as músicas que ele percebe, atua como uma memória perceptual de um modelo de canção e, em seguida, a imitá-lo, se do sexo masculino, ou é capaz de reconhecer e identificar parentes ou parceiro, se uma fêmea. Esta informação é armazenada no canora CVS é movido através de sua AR axonal outro núcleo, que será responsável pelo desenvolvimento de habilidades motoras na execução de cantar através do nervo hipoglosso craniano que abastece os músculos da siringe.
Há evidências de que estes locais são mais elevados nas Canárias, com uma música mais complexa em que é mais fácil, que aumenta de tamanho após a administração de testosterona e é maior durante a primavera, quando os níveis de testosterona são mais elevados, também é maior nos machos que nas fêmeas, ao contrário, diminui de tamanho após a fase reprodutiva, quando novamente a música torna-se instável e os níveis de testosterona no sangue cair acentuadamente.
Constatou-se também que a sobrevivência dos neurônios que chegam ao CVS recuperado depende dos níveis suficientes de testosterona, o que seria recrutar esses neurônios pelo derivado do cérebro em si fator neurotrófico (BDNF) mostrou que a sobrevivência depende do tempo cada rodada canário canta, como o uso de circuitos de motores associados com o canto estimula a liberação de BDNF, neste último caso, afeta mais neurônios no RAS central. Considerou também que a audição de sons ajuda a desenvolver e melhorar os circuitos auditivos envolvidos na aprendizagem da cantora.
A diminuição significativa nos níveis de testosterona durante os primeiros meses do primeiro ano de vida, de maio a julho, coincide com o aparecimento da primeira muda e estagnação no crescimento do tamanho da música drivers da escola, que é recuperada na meses, em paralelo com uma recuperação dos níveis de testosterona e é o mais elevado em novembro. Tudo isso ocorre durante a fase de plástico canção associada a um importante processo de aprendizagem. A dimensão destes centros cerebrais, como foi dito, é a maior durante a primavera, quando os níveis de testosterona são mais elevados.
A partir do segundo ano e anualmente no final da época de reprodução, durante a muda que vem, principalmente em agosto, os neurônios dos centros de regulação da canção são submetidos a regressão e morrer em grande número, causando uma diminuição no tamanho núcleos, coincidindo com uma diminuição significativa nos níveis de testosterona. Nesta fase de regressão dos núcleos é seguida por outra de regeneração, através de um processo de neurogênese com a incorporação de novos neurônios, que é máximo em novembro, quando os níveis de testosterona são mais elevados. Este processo é repetido a cada ano igual.
Coincidindo com este momento de crise para a queda e recuperação dos níveis de testosterona com regressão e regeneração dos condutores de núcleos cerebrais canção, há instabilidade e mudança no canto pode ficar perdido, adicionar ou alterar as sílabas da canção. A música torna-se mais estável entre novembro e maio, quando o processo de regeneração dos centros cerebrais está estabilizada. Estas flutuações no tamanho dos núcleos do cérebro não são vistos em outras aves canoras em que aprendeu a canção dura a vida inteira.
Dada a visita paralelismo entre a capacidade de aprender e entregar uma canção e tamanho cerebSuperposición núcleos complexos aprendeu o número de sílabas por mês, regulador de tamanho maior centro vocal (CVS) de cantar e os níveis de testosterona no sangue. Durante a canção de plástico ou "período crítico" de aprendizagem são consideradas frango baixos níveis de testosterona: após um declínio inicial é muito acentuada no primeiro semestre (maio-junho) é o "mute" e estagna agenda de crescimento para o CVS, então há uma recuperação gradual no segundo semestre (agosto a outubro), no segundo semestre, combinando um aumento dramático no tamanho do CVS pelo fenômeno da neurogênese, uma capacidade de aprendizagem e de recuperação dos níveis de testosterona , para adquirir níveis manimos em noviembre.rales, estrelando a ação da testosterona durante as fases de subcanción e principalmente durante a música de plástico, é refletida cantando dependência em relação ao correto desenvolvimento dos centros cerebrais e, por sua vez, a disponibilidade de adequados níveis de testosterona no momento.
Além disso, anualmente, em janeiro, há uma queda considerável dos níveis de testosterona no sangue de significado não muito claro, que não é acompanhado por uma instabilidade na canção ou diminuir o tamanho dos centros da música, talvez por um maior consumo desse hormônio nos tecidos para desenvolver as características sexuais em preparação para a temporada de reprodução
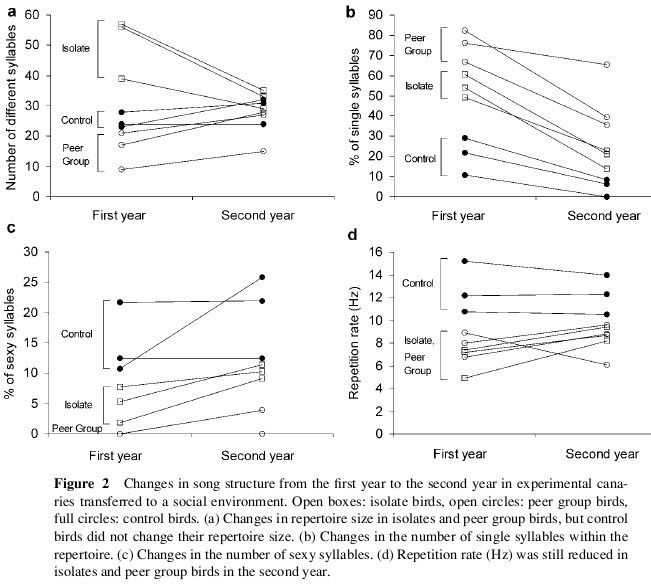
APRENDIZAGEM, núcleos e testosterona no cérebro do canário.
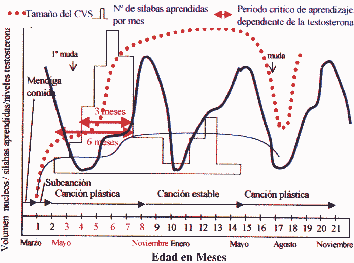
Cobertura do número de sílabas aprendidas por mês, regulador de tamanho maior centro vocal (CVS) de cantar e os níveis de testosterona no sangue. Durante a canção de plástico ou "período crítico" de aprendizagem são consideradas frango baixos níveis de testosterona: após um declínio inicial é muito acentuada no primeiro semestre (maio-junho) é o "mute" e estagna agenda de crescimento para o CVS, então há uma recuperação gradual no segundo semestre (agosto a outubro), no segundo semestre, combinando um aumento dramático no tamanho do CVS pelo fenômeno da neurogênese, uma capacidade de aprendizagem e de recuperação dos níveis de testosterona , para ganhar os níveis máximos em novembro.
No final da época de reprodução das Canárias como um adulto, a partir de Maio, retornando a níveis mais baixos de testosterona, é novamente o "mute" regressão do tamanho do CVS morte de seus neurônios e perda de especialização na performance da canção, voltando em fase de música de plástico. AI rali novamente os níveis de testosterona, isso induz a um novo processo de neurogênese, ele recupera o tamanho do CVS e em fase estável, com uma faixa de pontuação potencialmente diferentes para esquecer ou alterar notas antigas ou adquirir novos. Este ciclo é repetido a cada ano! É evidente dependência canção aprendizagem estável durante a canção, antes da Primavera, há outra queda significativa nos níveis de testosterona não tem impacto sobre o tamanho do CVS ou no domínio da música que, talvez, poderia estar em relação com o aumento do consumo do hormônio de preparar os tecidos do macho para a reprodução. (Modificado de Nottebohm F. De canto dos pássaros a neurogênese. Sci Am 1989 fevereiro: 260 / 2): 74-9)
APRENDIZAGEM E imitação de um modelo adulto.
É lógico que, se as Canárias são capazes de aprender a cantar imitando o modelo de machos adultos, e este é o procedimento a seguir quando estas aves vivem em liberdade na natureza, e vivendo e aprendendo o canto do pai e os outros machos adultos grupo nossa canários também devem aprender a cantar imitando um adulto do sexo masculino com uma música de qualidade, seguindo o exemplo da natureza. Este é o critério de bons criadores como Arcadio Macho Pavón, La Puebla de Cazalla (Sevilla), que usa o professor com as galinhas para que eles irão aprender a pronunciar as notas, desde que a genética lhes com uma boa voz. O professor deixou na "solta" ou gaiolas, junto com os filhotes, até que se deslocam para gaiolas individuais, os professores são canários muito completo que não tenham pisado ou levantadas em excesso, porque eles mantêm uma música de alta qualidade . Depois que os filhotes deixam o "solto" não é mais necessário pelo professor. A hipótese de que cantar é melhor quando a aprendizagem de uma cópia talentoso e evitar defeitos na música, previamente purificada professores.
Outros especialistas criadores como sandúa Joaquin Sanchez de San Pedro de Mieres (Asturias), argumentam, no entanto, a abordagem não utilizado para os professores de canto que exibem o frango é manifestação espontânea de seus genes, e não distorcer a aprendizagem expressão destes genes, portanto, a seleção que você está fazendo cópias dos melhores canções correspondem a amostras com um genótipo de maior qualidade. Isso não significa que dois canários com o mesmo fundo genético deve apresentar a mesma música como a canção, como mostra o fenótipo resulta da interação dos genes com o ambiente em seu sentido mais amplo (que sons audíveis, o nível de brilho, alimentação, temperatura, mais ou menos numerosos toques, o fornecimento de níveis corretos de testosterona durante a música de plástico, etc.) e é quase impossível que esse meio também é idêntica em ambos os pássaros. Este método de aprendizagem sem um professor seria aplicável, até que uma correção de rota em particular, então pode muito bem utilizar um professor ou não, uma vez que a seleção é feita ea linha fixa. Além disso, a experiência mostra-lhes que canário canção produzida uma pontuação ou nenhum professor tenha mais capacidade de improvisar, de modo que a música nunca se repete, mudando de uma vez para o outro. Seria útil para o grupo de galinhas para as famílias que têm um fundo genético similar e da canção deve ser de características semelhantes e evitar ou atenuar a influência de outros grupos de frangos de linhas diferentes, alojados em gaiolas adjacentes que poderiam ser ouvidas e se influenciam mutuamente ao que costumamos usar uma rádio que parece barreira acústica.
Uma abordagem intermediária, mais eclético, é o professor utilizando o "Loose", apenas até que ele começa a se mover em julho para parar de cantar os professores, assim que as galinhas poderiam desenvolver de acordo com sua canção genética própria aprendidas durante o resto do verão e cair. Uma quarta possibilidade, também usado por alguns criadores, é o emprego de vários professores com canções diferentes, tudo enfiado na "solta" e cada um pombo que iria aprender a música que você gosta ou uma mistura deles. Outros reprodutores usar um sistema misto: a introdução de novas floresce e canções novas, alguns pares raça distante sem professores e, se ele vem com alguma qualidade floresce novos ou novas canções, como professores de usá-lo no futuro.
Há estudos experimentais com outros pássaros, como o tentilhão, um dos quais é possível extrair as seguintes conclusões:
Um único pássaro criado em cativeiro com o seu pai quase completamente imita as músicas do último, mas quando vários frangos são criados em conjunto com um macho adulto, cada uma dessas galinhas tornam-se, transformar-se em tutores de seus irmãos e diminui a imitação do modelo de pai: quanto maior o número de galinhas em conjunto, o menor número de sílabas imitado e menor duração da canção aprendida. A tarde-nascidos filhotes adultos mais rápido do que os irmãos mais velhos, em termos relativos, e uma imitação mais completa.
Frangos aprender a partir de um modelo de canção por "playback" na gaiola, mas a superabundância deste modelo de imitação reduz até 33%. Esta restrição de imitação seletiva afeta sílabas específico, não ocorrem de forma aleatória, isto faz sílabas que quase todas as galinhas têm imitado e quase ninguém menos que o terno.
A criação de frangos com o isolamento social, ou seja, sem ouvir música sozinho e os padrões de um adulto pode levar a cantar de improviso, embora atípico, inclui muitos sons normais. No entanto, nem essa capacidade de improvisar parecem manifestar quando ela induz uma menor imitação de um modelo adulto, após a superexposição do mesmo.
Se as galinhas ouvir cantar um outro macho que está fora da gaiola, mas não conseguiu fazer contato visual ou interagir com ele, imitando a sua música será pouco ou nulo.
Não existem experiências, mas estão em andamento, que tenham usado os diversos padrões de música diferente simultaneamente.
À luz destes dados, e em relação aos hábitos descrito em nosso Canaricultura podemos dizer o seguinte:
Na verdade, as galinhas trancadas com a mãe ou outro adulto do sexo masculino canção imita a sua quase completa. A imitação é menos completo, tanto corresponder a mais galinhas em gaiolas de galinhas e mesmo do nascimento e falecido pai, aprendi a música tão cedo do que o mais velho. A utilização de gravações jogado na jaula imitação atinge quase completa, paradoxalmente, se as galinhas mal ouvi-los, não mais de 2-3 minutos por dia, se a reprodução é abundante, a imitação é menos completo.
Falha no uso de modelos adultos para imitação leva a improvisar uma canção frangos características atípicas, mas com sílabas. Neste caso, cada frango serviria como tutor de seus irmãos.
A utilização de um fundo de rádio, tocando todos os dias, pode produzir dois efeitos: primeiro, para estimular as vias auditivas da canção, incluindo o CVS, por outro, estimular superabundante auditivo, conseguindo reduzir a capacidade de imitação no frangos que ouvi-lo, se prosseguir o criador não é imitar outros modelos para aqueles que ouvem. Você pode imaginar a ação estimulante dos sons na via auditiva da canção, uma vez que, na ausência de estímulos, as vias são igualmente a atrofia como centros de pessoa surda atrofia e caminhos neurológicos da fala e nunca falar corretamente .
A exposição excessiva aos sons, como o uso de rádio de forma contínua ou aprendizagem em conjunto com um número significativo de frangos em conjunto, poderiam iniciar processos de seleção sob os cuidados de frangos de imitação, aceitando alguns e rejeitando outros modelos . Isso poderia explicar a rica diversidade de sons e músicas encontradas em pássaros, ambas as áreas naturais e em cativeiro.
CONCLUSÕES
Embora existam muitas questões relacionadas com a aprendizagem do canto canários, que estão abertos para estudo e experimentação, podemos estabelecer as seguintes conclusões:
As características do poder e do tom de voz, baseando-se principalmente na conformação anatômica do órgão fonatório o canário, são em grande parte determinado pelo genótipo.
A partitura musical ou a canário canção faz parte do fenótipo desta e, conseqüentemente, resulta da interação do genótipo com o ambiente em seu sentido mais amplo.
Apesar das Canárias mantêm a capacidade de aprender novas sílabas ao longo do ano, durante a música de plástico ou "período crítico" de aprendizagem quando elas têm mais capacidade e, conseqüentemente, o encurtamento deste período, a aceleração da maturação sexual por enjaular individual fornecimento cedo ou excesso de testosterona, a pobreza pode levar a uma sílaba da música.
Nós não sabemos muitos fatores ambientais que possam influenciar a aprendizagem do canto, como um aspecto específico do fenótipo, isto é, ser capaz de agrupá-los em dois: aqueles associados com estímulos auditivos que podem ou não podem servir de modelo e aqueles associados com a disponibilidade de ótimos níveis de testosterona.
É geralmente aceite que os professores não aprenderam uma canção com mais riqueza e sílaba mais musicalidade, embora seja claro que as galinhas cantar como os pais, dada a capacidade de imitação das pessoas. Aprender sem professor olha como ele poderia fazer uma música com pelo menos tanta riqueza quanto silábica e musicalidade com um professor, ajudá-los a alcançar novas e floresce músicas que são reflexo de um genótipo adequado, permitindo uma mais precisa seleção genética; alcançar uma maior capacidade risco de improvisação, mas não é maior que a canção é de pouco valor e deficiências na execução da canção. Não haveria nenhuma objeção ao uso de um professor em uma linha, quando este já foi fixado.
© Jerez Vicente Gómez-Coronado
OBS; Terá ainda que ser nalgumas partes redigido ,o que farei oportunamente .Isto é um trabalho minucioso pelo qual me guio pelo original ,isto foi-me enviado e mensagem privada e posteriormente postado o link no forum canaricultura de color ,pelo qual fiquei deveras satisfeito ,alguém reconheceu o meu ponto de análize mas cientifica .
Por acaso alguém comprou a Enciclopédia visual da Visão ?
Pois no de Aves vinha lá um artigo interessante sobre a comunicação das aves ,em geral trás bosn artigos ,a baixissimo custo ,1 euro
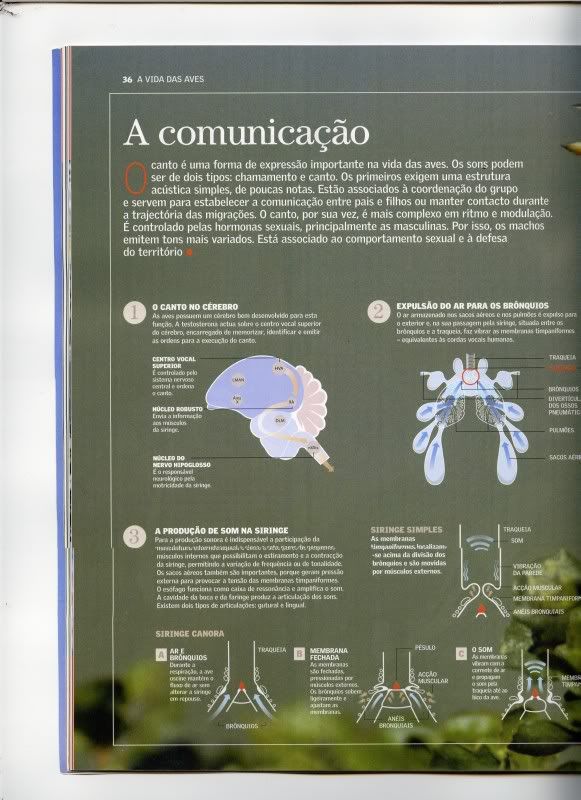

Se tiverem dificuldade em verem ,puxem a imagem pró vosso pc e visualizem a imagem ,utilizando a lupazunha ,vale bem a pena dar uma vista de olhos
Embora o canto canário é executado em seu órgão melodiosa, a siringe, é o cérebro onde reside o conteúdo no "Stable Song", com sua composição silábica e padrões específicos musical e quando a sua execução é controlada pela a siringe, como discutido abaixo.
Em 1976, Fernando Nottebohm, um especialista em comportamento animal e neurogênese de The Rockefeller University, NY, E.U.A., identificados grupos distintos de células cerebrais no controle da cantando canários. Estas células são distribuídos em um grande núcleo, chamado de "maior centro vocal (CVS), que envia axonal outro núcleo denominado" centro robustus archistriatalis (RA), esta, por sua vez, envia outros para o núcleo de axonal " nervo craniano hipoglosso "que abastece os músculos da siringe.
Cérebro Canárias. No desenho cima de um corte lateral do cérebro e na parte inferior do desenho de uma seção longitudinal da traquéia, brônquios e siringe do canário. O cérebro tem vários núcleos formados por neurônios especializados de controle de aprendizagem e performance da canção. Quanto maior o núcleo central superior vocal (CVS), que envia as extensões nervosas para o "núcleo Robustus archistriatalis" (M), que por sua vez, enviar outras extensões do nervo craniano ao núcleo do hipoglosso, que abastece os músculos da siringe: órgão onde o som é produzido. (Reproduzido de Nottebohm F. De canto dos pássaros a neurogênese. Sci Am 1989 Feb; 260 (2): 74-9).
Ambos os centros estão localizados nos lobos frontais das Canárias, que é onde você controla complexos comportamentos aprendidos. CVS parece ser o gerente da loja para identificar e produzir a música, na verdade, este kernel trabalha para reconhecer as músicas que ele percebe, atua como uma memória perceptual de um modelo de canção e, em seguida, a imitá-lo, se do sexo masculino, ou é capaz de reconhecer e identificar parentes ou parceiro, se uma fêmea. Esta informação é armazenada no canora CVS é movido através de sua AR axonal outro núcleo, que será responsável pelo desenvolvimento de habilidades motoras na execução de cantar através do nervo hipoglosso craniano que abastece os músculos da siringe.
Há evidências de que estes locais são mais elevados nas Canárias, com uma música mais complexa em que é mais fácil, que aumenta de tamanho após a administração de testosterona e é maior durante a primavera, quando os níveis de testosterona são mais elevados, também é maior nos machos que nas fêmeas, ao contrário, diminui de tamanho após a fase reprodutiva, quando novamente a música torna-se instável e os níveis de testosterona no sangue cair acentuadamente.
Constatou-se também que a sobrevivência dos neurônios que chegam ao CVS recuperado depende dos níveis suficientes de testosterona, o que seria recrutar esses neurônios pelo derivado do cérebro em si fator neurotrófico (BDNF) mostrou que a sobrevivência depende do tempo cada rodada canário canta, como o uso de circuitos de motores associados com o canto estimula a liberação de BDNF, neste último caso, afeta mais neurônios no RAS central. Considerou também que a audição de sons ajuda a desenvolver e melhorar os circuitos auditivos envolvidos na aprendizagem da cantora.
A diminuição significativa nos níveis de testosterona durante os primeiros meses do primeiro ano de vida, de maio a julho, coincide com o aparecimento da primeira muda e estagnação no crescimento do tamanho da música drivers da escola, que é recuperada na meses, em paralelo com uma recuperação dos níveis de testosterona e é o mais elevado em novembro. Tudo isso ocorre durante a fase de plástico canção associada a um importante processo de aprendizagem. A dimensão destes centros cerebrais, como foi dito, é a maior durante a primavera, quando os níveis de testosterona são mais elevados.
A partir do segundo ano e anualmente no final da época de reprodução, durante a muda que vem, principalmente em agosto, os neurônios dos centros de regulação da canção são submetidos a regressão e morrer em grande número, causando uma diminuição no tamanho núcleos, coincidindo com uma diminuição significativa nos níveis de testosterona. Nesta fase de regressão dos núcleos é seguida por outra de regeneração, através de um processo de neurogênese com a incorporação de novos neurônios, que é máximo em novembro, quando os níveis de testosterona são mais elevados. Este processo é repetido a cada ano igual.
Coincidindo com este momento de crise para a queda e recuperação dos níveis de testosterona com regressão e regeneração dos condutores de núcleos cerebrais canção, há instabilidade e mudança no canto pode ficar perdido, adicionar ou alterar as sílabas da canção. A música torna-se mais estável entre novembro e maio, quando o processo de regeneração dos centros cerebrais está estabilizada. Estas flutuações no tamanho dos núcleos do cérebro não são vistos em outras aves canoras em que aprendeu a canção dura a vida inteira.
Dada a visita paralelismo entre a capacidade de aprender e entregar uma canção e tamanho cerebSuperposición núcleos complexos aprendeu o número de sílabas por mês, regulador de tamanho maior centro vocal (CVS) de cantar e os níveis de testosterona no sangue. Durante a canção de plástico ou "período crítico" de aprendizagem são consideradas frango baixos níveis de testosterona: após um declínio inicial é muito acentuada no primeiro semestre (maio-junho) é o "mute" e estagna agenda de crescimento para o CVS, então há uma recuperação gradual no segundo semestre (agosto a outubro), no segundo semestre, combinando um aumento dramático no tamanho do CVS pelo fenômeno da neurogênese, uma capacidade de aprendizagem e de recuperação dos níveis de testosterona , para adquirir níveis manimos em noviembre.rales, estrelando a ação da testosterona durante as fases de subcanción e principalmente durante a música de plástico, é refletida cantando dependência em relação ao correto desenvolvimento dos centros cerebrais e, por sua vez, a disponibilidade de adequados níveis de testosterona no momento.
Além disso, anualmente, em janeiro, há uma queda considerável dos níveis de testosterona no sangue de significado não muito claro, que não é acompanhado por uma instabilidade na canção ou diminuir o tamanho dos centros da música, talvez por um maior consumo desse hormônio nos tecidos para desenvolver as características sexuais em preparação para a temporada de reprodução
APRENDIZAGEM, núcleos e testosterona no cérebro do canário.
Cobertura do número de sílabas aprendidas por mês, regulador de tamanho maior centro vocal (CVS) de cantar e os níveis de testosterona no sangue. Durante a canção de plástico ou "período crítico" de aprendizagem são consideradas frango baixos níveis de testosterona: após um declínio inicial é muito acentuada no primeiro semestre (maio-junho) é o "mute" e estagna agenda de crescimento para o CVS, então há uma recuperação gradual no segundo semestre (agosto a outubro), no segundo semestre, combinando um aumento dramático no tamanho do CVS pelo fenômeno da neurogênese, uma capacidade de aprendizagem e de recuperação dos níveis de testosterona , para ganhar os níveis máximos em novembro.
No final da época de reprodução das Canárias como um adulto, a partir de Maio, retornando a níveis mais baixos de testosterona, é novamente o "mute" regressão do tamanho do CVS morte de seus neurônios e perda de especialização na performance da canção, voltando em fase de música de plástico. AI rali novamente os níveis de testosterona, isso induz a um novo processo de neurogênese, ele recupera o tamanho do CVS e em fase estável, com uma faixa de pontuação potencialmente diferentes para esquecer ou alterar notas antigas ou adquirir novos. Este ciclo é repetido a cada ano! É evidente dependência canção aprendizagem estável durante a canção, antes da Primavera, há outra queda significativa nos níveis de testosterona não tem impacto sobre o tamanho do CVS ou no domínio da música que, talvez, poderia estar em relação com o aumento do consumo do hormônio de preparar os tecidos do macho para a reprodução. (Modificado de Nottebohm F. De canto dos pássaros a neurogênese. Sci Am 1989 fevereiro: 260 / 2): 74-9)
APRENDIZAGEM E imitação de um modelo adulto.
É lógico que, se as Canárias são capazes de aprender a cantar imitando o modelo de machos adultos, e este é o procedimento a seguir quando estas aves vivem em liberdade na natureza, e vivendo e aprendendo o canto do pai e os outros machos adultos grupo nossa canários também devem aprender a cantar imitando um adulto do sexo masculino com uma música de qualidade, seguindo o exemplo da natureza. Este é o critério de bons criadores como Arcadio Macho Pavón, La Puebla de Cazalla (Sevilla), que usa o professor com as galinhas para que eles irão aprender a pronunciar as notas, desde que a genética lhes com uma boa voz. O professor deixou na "solta" ou gaiolas, junto com os filhotes, até que se deslocam para gaiolas individuais, os professores são canários muito completo que não tenham pisado ou levantadas em excesso, porque eles mantêm uma música de alta qualidade . Depois que os filhotes deixam o "solto" não é mais necessário pelo professor. A hipótese de que cantar é melhor quando a aprendizagem de uma cópia talentoso e evitar defeitos na música, previamente purificada professores.
Outros especialistas criadores como sandúa Joaquin Sanchez de San Pedro de Mieres (Asturias), argumentam, no entanto, a abordagem não utilizado para os professores de canto que exibem o frango é manifestação espontânea de seus genes, e não distorcer a aprendizagem expressão destes genes, portanto, a seleção que você está fazendo cópias dos melhores canções correspondem a amostras com um genótipo de maior qualidade. Isso não significa que dois canários com o mesmo fundo genético deve apresentar a mesma música como a canção, como mostra o fenótipo resulta da interação dos genes com o ambiente em seu sentido mais amplo (que sons audíveis, o nível de brilho, alimentação, temperatura, mais ou menos numerosos toques, o fornecimento de níveis corretos de testosterona durante a música de plástico, etc.) e é quase impossível que esse meio também é idêntica em ambos os pássaros. Este método de aprendizagem sem um professor seria aplicável, até que uma correção de rota em particular, então pode muito bem utilizar um professor ou não, uma vez que a seleção é feita ea linha fixa. Além disso, a experiência mostra-lhes que canário canção produzida uma pontuação ou nenhum professor tenha mais capacidade de improvisar, de modo que a música nunca se repete, mudando de uma vez para o outro. Seria útil para o grupo de galinhas para as famílias que têm um fundo genético similar e da canção deve ser de características semelhantes e evitar ou atenuar a influência de outros grupos de frangos de linhas diferentes, alojados em gaiolas adjacentes que poderiam ser ouvidas e se influenciam mutuamente ao que costumamos usar uma rádio que parece barreira acústica.
Uma abordagem intermediária, mais eclético, é o professor utilizando o "Loose", apenas até que ele começa a se mover em julho para parar de cantar os professores, assim que as galinhas poderiam desenvolver de acordo com sua canção genética própria aprendidas durante o resto do verão e cair. Uma quarta possibilidade, também usado por alguns criadores, é o emprego de vários professores com canções diferentes, tudo enfiado na "solta" e cada um pombo que iria aprender a música que você gosta ou uma mistura deles. Outros reprodutores usar um sistema misto: a introdução de novas floresce e canções novas, alguns pares raça distante sem professores e, se ele vem com alguma qualidade floresce novos ou novas canções, como professores de usá-lo no futuro.
Há estudos experimentais com outros pássaros, como o tentilhão, um dos quais é possível extrair as seguintes conclusões:
Um único pássaro criado em cativeiro com o seu pai quase completamente imita as músicas do último, mas quando vários frangos são criados em conjunto com um macho adulto, cada uma dessas galinhas tornam-se, transformar-se em tutores de seus irmãos e diminui a imitação do modelo de pai: quanto maior o número de galinhas em conjunto, o menor número de sílabas imitado e menor duração da canção aprendida. A tarde-nascidos filhotes adultos mais rápido do que os irmãos mais velhos, em termos relativos, e uma imitação mais completa.
Frangos aprender a partir de um modelo de canção por "playback" na gaiola, mas a superabundância deste modelo de imitação reduz até 33%. Esta restrição de imitação seletiva afeta sílabas específico, não ocorrem de forma aleatória, isto faz sílabas que quase todas as galinhas têm imitado e quase ninguém menos que o terno.
A criação de frangos com o isolamento social, ou seja, sem ouvir música sozinho e os padrões de um adulto pode levar a cantar de improviso, embora atípico, inclui muitos sons normais. No entanto, nem essa capacidade de improvisar parecem manifestar quando ela induz uma menor imitação de um modelo adulto, após a superexposição do mesmo.
Se as galinhas ouvir cantar um outro macho que está fora da gaiola, mas não conseguiu fazer contato visual ou interagir com ele, imitando a sua música será pouco ou nulo.
Não existem experiências, mas estão em andamento, que tenham usado os diversos padrões de música diferente simultaneamente.
À luz destes dados, e em relação aos hábitos descrito em nosso Canaricultura podemos dizer o seguinte:
Na verdade, as galinhas trancadas com a mãe ou outro adulto do sexo masculino canção imita a sua quase completa. A imitação é menos completo, tanto corresponder a mais galinhas em gaiolas de galinhas e mesmo do nascimento e falecido pai, aprendi a música tão cedo do que o mais velho. A utilização de gravações jogado na jaula imitação atinge quase completa, paradoxalmente, se as galinhas mal ouvi-los, não mais de 2-3 minutos por dia, se a reprodução é abundante, a imitação é menos completo.
Falha no uso de modelos adultos para imitação leva a improvisar uma canção frangos características atípicas, mas com sílabas. Neste caso, cada frango serviria como tutor de seus irmãos.
A utilização de um fundo de rádio, tocando todos os dias, pode produzir dois efeitos: primeiro, para estimular as vias auditivas da canção, incluindo o CVS, por outro, estimular superabundante auditivo, conseguindo reduzir a capacidade de imitação no frangos que ouvi-lo, se prosseguir o criador não é imitar outros modelos para aqueles que ouvem. Você pode imaginar a ação estimulante dos sons na via auditiva da canção, uma vez que, na ausência de estímulos, as vias são igualmente a atrofia como centros de pessoa surda atrofia e caminhos neurológicos da fala e nunca falar corretamente .
A exposição excessiva aos sons, como o uso de rádio de forma contínua ou aprendizagem em conjunto com um número significativo de frangos em conjunto, poderiam iniciar processos de seleção sob os cuidados de frangos de imitação, aceitando alguns e rejeitando outros modelos . Isso poderia explicar a rica diversidade de sons e músicas encontradas em pássaros, ambas as áreas naturais e em cativeiro.
CONCLUSÕES
Embora existam muitas questões relacionadas com a aprendizagem do canto canários, que estão abertos para estudo e experimentação, podemos estabelecer as seguintes conclusões:
As características do poder e do tom de voz, baseando-se principalmente na conformação anatômica do órgão fonatório o canário, são em grande parte determinado pelo genótipo.
A partitura musical ou a canário canção faz parte do fenótipo desta e, conseqüentemente, resulta da interação do genótipo com o ambiente em seu sentido mais amplo.
Apesar das Canárias mantêm a capacidade de aprender novas sílabas ao longo do ano, durante a música de plástico ou "período crítico" de aprendizagem quando elas têm mais capacidade e, conseqüentemente, o encurtamento deste período, a aceleração da maturação sexual por enjaular individual fornecimento cedo ou excesso de testosterona, a pobreza pode levar a uma sílaba da música.
Nós não sabemos muitos fatores ambientais que possam influenciar a aprendizagem do canto, como um aspecto específico do fenótipo, isto é, ser capaz de agrupá-los em dois: aqueles associados com estímulos auditivos que podem ou não podem servir de modelo e aqueles associados com a disponibilidade de ótimos níveis de testosterona.
É geralmente aceite que os professores não aprenderam uma canção com mais riqueza e sílaba mais musicalidade, embora seja claro que as galinhas cantar como os pais, dada a capacidade de imitação das pessoas. Aprender sem professor olha como ele poderia fazer uma música com pelo menos tanta riqueza quanto silábica e musicalidade com um professor, ajudá-los a alcançar novas e floresce músicas que são reflexo de um genótipo adequado, permitindo uma mais precisa seleção genética; alcançar uma maior capacidade risco de improvisação, mas não é maior que a canção é de pouco valor e deficiências na execução da canção. Não haveria nenhuma objeção ao uso de um professor em uma linha, quando este já foi fixado.
© Jerez Vicente Gómez-Coronado
OBS; Terá ainda que ser nalgumas partes redigido ,o que farei oportunamente .Isto é um trabalho minucioso pelo qual me guio pelo original ,isto foi-me enviado e mensagem privada e posteriormente postado o link no forum canaricultura de color ,pelo qual fiquei deveras satisfeito ,alguém reconheceu o meu ponto de análize mas cientifica .
Por acaso alguém comprou a Enciclopédia visual da Visão ?
Pois no de Aves vinha lá um artigo interessante sobre a comunicação das aves ,em geral trás bosn artigos ,a baixissimo custo ,1 euro
Se tiverem dificuldade em verem ,puxem a imagem pró vosso pc e visualizem a imagem ,utilizando a lupazunha ,vale bem a pena dar uma vista de olhos



")